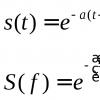Рефлекс и рефлекторная деятельность нервной системы. Рефлекторный принцип деятельности нервной системы, целостного организма. Рефлекторный принцип деятельности ЦНС
Взаимодействие нервных клеток составляет основу целенаправленной деятельности нервной системы и прежде всего осуществления рефлекторных актов. Таким образом, нервная регуляция носит рефлекторный характер.
Рефлексом называют ответную реакцию организма на раздражение рецепторов, осуществляемую через центральную нервную систему (ЦНС). Основные положения рефлекторного принципа деятельности ЦНС разрабатывались на протяжении двух с половиной столетий. Ученые выделяют пять этапов развития данной концепции.
Первый этап . Связан с формированием в ХУ11 столетии основ понимания рефлекторного принципа деятельности ЦНС. Принцип рефлекторной (отражательной) деятельности нервной системы был выдвинут в 17 столетии французским философом и математиком Рене Декартом, который считал, что все вещи и явления можно объяснить естественнонаучным путем. Данная исходная позиция позволила Р.Декарту сформулировать два важных положения рефлекторной теории:
1) деятельность организма при внешнем воздействии является отраженной (впоследствии ее стали называть рефлекторной – от лат reflexus – отраженный);
2) ответная реакция на раздражение осуществляется при помощи нервной системы.
Согласно теории Р.Декарта, нервы – это трубочки, по которым с огромной скоростью движутся животные духи, материальные частицы неизвестной природы. По нервам они попадают в мышцу, которая в результате раздувается (сокращается).
Второй этап . Связан с экспериментальным обоснованием материалистических представлений о рефлексе (ХУ11 – ХУ111 вв.). В частности, было установлено, что рефлекторная реакция может осуществляться на одном метамере лягушки (метаме р - сегмент спинного мозга, связанный с «кусочком тела»). Значительный вклад в развитие представлений о рефлекторной деятельности нервной системы внес чешский физиолог18 столетия И.Прохазка, который исходил из признания единства организма и окружающей среды, а также утверждал ведущую роль нервной системы в регуляции функций организма. Именно И.Прохазкой был предложен сам же термин «рефлекс». Кроме того, он ввел в физиологию закон силы (увеличение силы стимула увеличивает силу рефлекторной реакции организма; стимулы имеются не только внешние, но и внутренние); впервые дал описание классической рефлекторной дуги. В этот временной отрезок учеными в результате клинических экспериментальных исследований установлена роль задних (чувствительных) и передних (двигательных) корешков спинного мозга (закон Белла – Мажанди). Активно рефлекторная деятельность (в частности, сегментарные рефлексы) изучает Ч.Шеррингтон. В результате своих научных изысканий ученый описывает принцип афферентной иннервации мышц-антагонистов, вводит понятие «синапс», принцип общего нервного пути, понятие об интегративной деятельности нервной системы.
Третий этап . Утверждаются материалистические представления о психической деятельности (И.М.Сеченов, 60-е годы Х1Х в). Наблюдая за развитием детей, ученый приходит к выводу о том, что в основе формирования психической деятельности лежит именно принцип рефлекса. Это свое утверждение он выразил в следующей фрезе: «Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». При изучении рефлексов он обосновал приспособительный характер изменчивости рефлекса, открыл механизм торможения рефлексов, а также механизм суммации возбуждения в ЦНС.
Четвертый этап . Связан с разработкой основ учения о высшей нервной деятельности (исследования И.П.Павлова, начало ХХ столетия). И.П.Павлов открыл условные рефлексы и использовал их как объективный метод в изучении психической деятельности (высшей нервной деятельности). Ученым были сформулированы три основных принципа рефлекторной теории:
1. Принцип детерминизма (принцип причинности), согласно которому любая рефлекторная реакция причинно обусловлена. И.П.Павлов утверждал: «Нет действия без причины». Всякая деятельность организма, каждый акт нервной деятельности вызван определенной причиной, воздействием из внешнего мира или внутренней среды организма. Целесообразность реакции определяется специфичностью раздражителя, чувствительностью к ним (раздражителям) организма.
2. Принцип структурности. Его сущность заключается в том, что рефлекторная реакция осуществляется с помощью определенных структур. Чем больше структур, структурных элементов участвует в осуществлении этой реакции, тем она совершеннее. В мозге нет процессов, которые не имели бы материальной основы. Каждый физиологический акт нервной деятельности приурочен к определенной структуре.
3. Принцип единства процессов анализа и синтеза в составе рефлекторной реакции. Нервная система анализирует, т.е. различает, с помощью рецепторов все действующие внешние и внутренние раздражители и на основании этого анализа формирует целостную ответную реакцию – синтез. Анализ и синтез как поступающей информации, так и ответных реакций происходит в мозге непрерывно. В результате организм извлекает из окружающей среды полезную информацию, перерабатывает ее, фиксирует в памяти и формирует ответные действия в соответствии с обстоятельствами и потребностями.
Пятый этап . Характеризуется созданием учения о функциональных системах (исследования П.К.Анохина, середина ХХ в). Функциональная система – это динамическая совокупность различных органов и тканей, формирующаяся для достижения полезного (приспособительного) результата. Полезным результатом является поддержание постоянства внутренней среды организма с помощью регуляции функций внутренних органов и поведенческой соматической регуляции (например, поиск и потребление воды при ее недостатке в организме и возникновении жажды – биологической потребности). Полезным результатом может быть и удовлетворение социальной потребности (достижение высоких результатов учебной деятельности).
Исследуя рефлекторную основу жизнедеятельности живых организмов, ученые пришли к выводу о том, что базовыми являются рефлексы врожденные (безусловные), поскольку именно эти рефлексы, сформировавшиеся за миллионы лет эволюции, одинаковы для всех представителей конкретного вида животных организмов и мало зависят от ситуативных условий существования того или иного конкретного представителя данного вида животных. При резком же изменении условий окружающей среды безусловный рефлекс может привести и к гибели организма.
Безусловные рефлексы – ответная реакция организма на раздражение сенсорных рецепторов, осуществляемая с помощью нервной системы. И.П.Павлов выделил, в первую очередь, безусловные рефлексы, направленные на самосохранение организма (основными здесь являются пищевые, оборонительные, ориентировочные и некоторые другие). Данные рефлексы составляют большие группы разнообразных врожденных реакций.
Безусловно-рефлекторная деятельность изучалась П.В.Сомоновым. По мнению ученого, освоению каждой сферы среды соответствуют три разных класса безусловных рефлексов:
· витальные безусловные рефлексы, которые обеспечивают индивидуальное и видовое сохранение организма (пищевой, питьевой, регуляция сна, оборонительный и ориентировочный, рефлекс экономии сил и т.д.). Критериями данных рефлексов являются: физическая гибель особи в результате неудовлетворения соответствующей потребности, реализация безусловного рефлекса без участия другой особи того же вида;
· ролевые (зоосоциальные). Могут быть реализованы лишь только путем взаимодействия с другими особями своего вида. Данные рефлексы лежат в основе территориального, родительского и т.п. поведения. Кроме того, они имеют огромное значение для феномена эмоционального резонанса, «сопереживания» и формирования групповой иерархии, где каждая отдельная особь неизменно выступает в той или иной роли (брачного партнера, родителя или детеныша, хозяина территории или пришельца, лидера или ведомого и т.д.);
· безусловные рефлексы саморазвития. Они ориентированы на освоение новых пространственно-временных сред, обращены к будущему. К их числу относится исследовательское поведение, безусловный рефлекс сопротивления (свободы), имитационный (подражательный) и игровой.
К числу безусловных рефлексов ученые относят и ориентировочный рефлекс. Ориентировочный рефлекс – безусловно-рефлекторное непроизвольное сенсорное внимание, сопровождаемое повышением тонуса мышц, вызванное неожиданным или новым для организма раздражителем. Данную реакцию ученые часто называют рефлексом настораживания, тревоги, удивления, а И.П.Павлов определил ее как рефлекс «что такое?». Ориентировочный рефлекс характеризуется проявлением целого комплекса реакций. Ученые выделяют три фазы в развитии данного рефлекса.
Первая фаза. Характеризуется прекращением текущей деятельности и фиксацией позы. По мнению П.В.Симонова, это общее (превентивное) торможение, которое возникает на появление любого постороннего раздражителя с неизвестным сигнальным значением.
Вторая фаза . Она начинается, когда состояние «стоп-реакции» переходит в реакцию активации. На данной фазе весь организм переводится в состояние рефлекторной готовности к возможной встрече с чрезвычайной ситуацией, что проявляется, выражается в общем повышении тонуса всей скелетной мускулатуры. На данной фазе ориентировочный рефлекс проявляется в форме поликомпонентной реакции, включающей в себя поворот головы и глаз в направлении стимула.
Третья фаза . Начинается с фиксации поля раздражителя для развертывания процесса дифференцированного анализа внешних сигналов и принятия решения об ответной реакции организма.
Поликомпонентный состав ориентировочного рефлекса свидетельствует о его сложной морфофункциональной организации.
Ориентировочный рефлекс входит в структуру ориентировочного поведения (ориентировочно-исследовательской деятельности), что особенно ярко проявляется в новой обстановке. Исследовательская деятельность здесь может быть направлена как на освоение новизны, удовлетворение любопытства, так и на поиск раздражителя, объекта, способного удовлетворить эту потребность. Кроме того, ориентировочный рефлекс направлен и на определение «значимости» раздражителя. При этом наблюдается повышение чувствительности анализаторов, что облегчает восприятие воздействующих на организм раздражителей и определение их значения.
Механизм осуществления ориентировочного рефлекса является результатом динамического взаимодействия между множеством различных образований специфических и неспецифических систем ЦНС. Так, фазу общей активации связывают главным образом с активацией стволовой ретикулярной формации и генерализованным возбуждением коры. В развитии фазы анализа стимула ведущее место занимает корково-лимбико-таламическая интеграция. При этом важную роль играет гиппокамп. Это обеспечивает специализированность процессов анализа «новизны» и «значимости» стимула.
Наряду с безусловными рефлексами которые могут быть отнесены к низшей нервной деятельности, у высших животных и человека на основе этой низшей нервной деятельности сформировались новые механизмы приспособления к постоянно меняющимся условиям окружающей среды – высшая нервная деятельность. С ее помощью, а конкретнее, при помощи условных рефлексов, эти живые организмы приобрели способность реагировать не только на непосредственное воздействие биологически значимых агентов (пищевых, оборонительных и пр.), но и на их отдаленные признаки.
На рубеже Х1Х и ХХ столетий известный русский физиолог И.П.Павлов, долгое время изучавший функции пищеварительных желез (за эти исследования ученый был удостоен Нобелевской премии в 1904г.), обнаружил у экспериментальных животных регулярное повышение секреции слюны и желудочного сока не только при попадании пищи в ротовую полость, а затем в желудок, но и при одном лишь ожидании приема пищи. В то время механизм такого явления был неизвестен и объяснялся «психическим возбуждением слюнных желез». В результате дальнейших научных исследований в данном направлении это явление ученым было названо как условные рефлексы . По мнению И.П.Павлова, условные рефлексы вырабатываются на основе безусловных и являются приобретенными в процессе жизнедеятельности. Кроме того, условные рефлексы непостоянны, то есть, могут появляться и исчезать на протяжении жизни человека в зависимости от меняющихся условий существования. Приобретение условных рефлексов происходит на протяжении всей жизни человека. Оно обусловлено непосредственным, постоянно изменяющимся окружением. Вновь приобретенные условные рефлексы многократно увеличивают и расширяют диапазон приспособительных реакций животных и человека.
Для выработки условного рефлекса необходимо совпадение во времени двух раздражителей, действующих на животное (или человека). Один из этих раздражителей при любых обстоятельствах вызывает закономерную рефлекторную реакцию, классифицируемую как безусловный рефлекс. Сам же такой раздражитель определяется как рефлекс условный. Другой раздражитель, применяемый для выработки условного рефлекса, в силу своей обыденности, как правило, не вызывает никакой реакции и определяется как индифферентный (безразличный). Раздражители такого рода лишь только при первых предъявлениях вызывают определенную ответную ориентировочную реакцию, которая, например, может проявляться в повороте головы и глаз в сторону действующего стимула. При повторных действиях стимула (раздражителя) ориентировочный рефлекс ослабевает, а затем и полностью исчезает в результате механизма привыкания, и тогда вызвавший его раздражитель становится индифферентным.
Как показали многочисленные исследования И.П.Павлова и его коллег, условный рефлекс вырабатывается при соблюдении следующих правил:
1. Индифферентный раздражитель должен подействовать на несколько секунд раньше, чем безусловный стимул. Исследования И.П.Павлова, проводимые на собаках, показали, что если, например, индифферентный раздражитель (различные звуковые сигналы) начнет действовать непосредственно в процессе кормления, а не до его начала, то условный рефлекс не образуется.
2. Биологическая значимость индифферентного раздражителя должна быть меньшей, чем у безусловного раздражителя. Опять же, ссылаясь на проводимые в лаборатории И.П.Павлова исследования, необходимо отметить, что если, например, использовать слишком громкие, пугающие звуковые сигналы, давая животному сразу после этого пищу, условный рефлекс не образуется.
3. Образованию условного рефлекса не должны мешать посторонние раздражители, отвлекающие на себя внимание животного.
О выработанном условном рефлексе можно говорить в том случае, если ранее индифферентный стимул начнет вызывать такую же реакцию, как и безусловный раздражитель, используемый в сочетании с ним. Так, если кормлению животного несколько раз предшествовало включение какого-либо звукового сигнала и в результате этого сочетания в дальнейшем только лишь при звуковом сигнале стало возникать слюноотделение, то эту реакцию следует считать проявлением условного рефлекса. Действие безусловного раздражителя вслед за индифферентным определяется как подкрепление, а когда индифферентный прежде стимул начинает вызывать рефлекторную реакцию, он становится условным раздражителем (условным сигналом).
Существует несколько подходов к классификации условных рефлексов.
В первую очередь, все условные рефлексы ученые делят (как и безусловные) на следующие группы.
По биологическому значению их различают на пищевые, оборонительные и т.д.
По виду рецепторов , с которых идет выработка, условные рефлексы делят на экстерорецептивные, проприорецептивные, интерорецептивные. В исследованиях В.М.Быкова и В.Н.Черниговского с их коллегами была показана связь коры больших полушарий со всеми внутренними органами. Интерорецептивные условные рефлексы сопровождаются, как правило, расплывчатыми ощущениями, которые еще И.М.Сеченов определил как «темные чувства», влияющие на настроение и работоспособность. Проприорецептивные условные рефлексы лежат в основе научения двигательным навыкам (ходьбе, производственным операциям и пр.). Экстерорецептивные условные рефлексы формируют приспособительное поведение животных по добыванию пищи, избеганию вредных воздействий, продолжению рода и т.д. Для человека важнейшее значение имеют экстерорецептивные словесные раздражители, формирующие поступки и мысли.
По функции отдела нервной системы и характеру эфферентного ответа различают условные рефлексы соматические (двигательные) и вегетативные (сердечно-сосудистые, секреторные, выделительные и пр.).
По отношению сигнального раздражителя к безусловному (подкрепляющему) раздражителю все условные рефлексы делят на натуральные и искусственные (лабораторные). Натуральные условные рефлексы формируются на сигналы, являющиеся естественными признаками подкрепляющего раздражителя (запах¸ цвет, определенное время и т.д.). Например, прием пищи в одно и то же время ведет к выделению пищеварительных соков и некоторых других реакций организма (например, лейкоцитоз к моменту приема пищи). Искусственными (лабораторными) называют условные рефлексы на такие сигнальные раздражители, которые в природе не имеют отношения к безусловному (подкрепляющемуся) раздражителю. Основными из этих условных рефлексов являются следующие:
· по сложности различают: простые условные рефлексы, вырабатываемые на одиночные раздражители (классические условные рефлексы, открытые И.П.Павловым); комплексные условные рефлексы (рефлексы, образующиеся на воздействие нескольких сигналов, действующих либо одновременно, либо последовательно); цепные рефлексы – рефлексы на цепь раздражителей, каждый из которых вызывает свой условный рефлекс (типичным примером здесь может быть динамический стереотип),
· по соотношению времени действия условного и безусловного раздражителей различают рефлексы наличные и следовые. Для выработки условных наличных рефлексов характерно совпадение действия условного и безусловного раздражителей. Следовые рефлексы вырабатываются в условиях, когда безусловный раздражитель подключается несколько позднее по времени (через 2-3 мин), чем условный. Т.Е. выработка условного рефлекса происходит на след сигнального стимула,
· по выработке условного рефлекса на базе другого условного рефлекса различают условные рефлексы первого, второго, третьего и других порядков. Рефлексы первого порядка – это условные рефлексы, выработанные на базе безусловных рефлексов (классические условные рефлексы). Рефлексы второго порядка вырабатываются на базе условных рефлексов первого порядка, при которых безусловный стимул отсутствует. Рефлекс третьего порядка образуется на базе рефлекса второго порядка и т.д. Чем выше порядок условного рефлекса, тем труднее идут его выработка. Так, у собак удается выработать лишь условные рефлексы третьего порядка (не выше),
· условные рефлексы на время могут быть не только натуральными, но и искусственными. При неоднократной подаче безусловного раздражителя с неизменным интервалом между подачами образуется рефлекс на время. Т.Е.за некоторое время до подачи подкрепления возникает условная эффекторная реакция.
В зависимости от сигнальной системы различают условные рефлексы на сигналы первой и второй сигнальных систем, т.е. на внешнее воздействие и на речь.
Кроме того, условные рефлексы могут быть положительными и отрицательными .
Многие ученые определяют условные рефлексы как реакции на будущие события. Биологическое значение условных рефлексов состоит в их предупредительной роли. Для организма они имеют приспособительное значение, готовя организм к будущей полезной поведенческой деятельности и помогая ему избежать вредных воздействий, тонко и эффективно адаптироваться к окружающей природной и социальной среде. Также необходимо отметить, что условные рефлексы формируются благодаря пластичности нервной системы.
Общая характеристика безусловных и условных рефлексов представлена в Таблице 1.
Таблице 1
Общая характеристика безусловных и условных рефлексов
Путь, по которому распространяется возбуждение при осуществлении рефлекса называют рефлекторной дугой (Рис 2).
Рефлекторная дуга состоит из пяти основных звеньев:
1. Рецептор.
2. Чувствительный путь.
3. Центральная нервная система.
4. Двигательный путь.
5. Рабочий орган.
Рис.2. Рефлекторная дуга:
а – двухнейронная; б – трехнейртонная
1 – рецептор; 2 – чувствительный (центростремительный) нерв; 3 – чувствительный нейрон в спинномозговой глии; 4 – аксон чувствительного нейрона; 5 – задние корешки спинномозговых нервов; 6 – вставочный нейрон; 7 – аксон вставочного нерва; 8 – двигательный нейрон в рогах спинного мозга; 9 – спинной мозг; 10 – аксон двигательного (центробежного) нейрона; 11 – рабочий орган.
Рефлекторная дуга представляет собой цепь нервных клеток, включающую афферентный (чувствительный) и эффекторный (двигательный, или секреторный) нейроны, по которым нервный импульс движется от места своего возникновения (от рецептора) к рабочему органу (эффектору). Большинство рефлексов осуществляется при участии рефлекторных дуг, которые образованы нейронами низших отделов ЦНС – нейронами спинного мозга.
Простейшая рефлекторная дуга состоит только из двух нейронов – афферентного (рецепторного) и эффекторного (эфферентного). Тело первого нейрона (афферентного), находится вне ЦНС. Как правило, это так называемый униполярный нейрон, тело которого расположено в спинномозговом узле или в чувствительном узле черепных нервов. Периферический отросток этой клетки находится в составе спинномозговых нервов или имеющих чувствительные волокна черепных нервов и их ветвей и заканчивается рецептором, воспринимающим внешнее (из внешней среды) или внутреннее (в органах, тканях организма) раздражение. Данное раздражение трансформируется рецептором в нервный импульс, который достигает тела нервной клетки, а затем по центральному отростку (совокупность таких отростков образует задние, чувствительные корешки спинномозговых нервов) направляется в спинной мозг или по соответствующим черепным нервам – в головной мозг. В сером веществе спинного мозга или в двигательном ядре головного мозга этот отросток чувствительной клетки образует синапс с толом второго нейрона (эфферентного). В межнейронном синапсе при помощи медиаторов происходит передача нервного возбуждения с чувствительного (афферентного) нейрона на двигательный (эфферентный) нейрон, отросток которого выходит из спинного мозга в составе передних корешков спинномозговых нервов или двигательных (секреторных) нервных волокон черепных нервов и направляется к рабочему органу, вызывая сокращение мышц, или торможение, или усиление секреции железы.
Сложная рефлекторная дуга . Как правило, рефлекторная дуга состоит не из двух нейронов и устроена гораздо сложнее. Между двумя нейронами – рецепторным (афферентным) и эффекторным (эфферентным) – имеется один или несколько замыкательных (вставочных) нейронов. В этом случае возбуждение от рецепторного нейрона по его центральному отростку передается не прямо эффекторной нервной клетке, а одному или нескольким вставочным нейронам. Роль вставочных нейронов в спинном мозге выполняют клетки, расположенные в сером веществе задних столбов. Часть этих клеток имеет аксон (нейрит), который направляется к двигательным клеткам передних рогов спинного мозга того же уровня и замыкает рефлекторную дугу на уровне данного сегмента спинного мозга. Аксон других клеток может в спинном мозге предварительно Т-образно делиться на нисходящую и восходящую ветви, которые направляются к двигательным клеткам передних рогов соседних, выше- и нижележащих сегментов. На пути следования каждая из отмеченных восходящих или нисходящих ветвей может отдавать коллатерали к двигательным клеткам этих и других соседних сегментов. В связи с этим, необходимо отметить, что раздражение даже самого минимального числа рецепторов может передаваться не только нервным клеткам какого-то определенного сегмента спинного мозга, но и распространяться на клетки нескольких соседних сегментов. В результате ответная реакция представляет собой сокращение не одной мышцы или одной группы мышц, а сразу нескольких групп. Таким образом, в ответ на раздражение возникает сложное рефлекторное движение – рефлекс.
Как мы отмечали выше, И.М.Сеченов в своем труде «Рефлексы головного мозга» выдвинул идею причинности (детерминизма), отмечая, что каждое явление в организме имеет свою причину, и рефлекторный эффект является ответом на эту причину. Эти идеи получили продолжение и подтверждение в трудах И.П.Павлова и С.П.Боткина. Именно И.П.Павлов распространил учение о рефлексе на всю нервную систему, начиная от ее низших отделов и кончая высшими ее отделами, и экспериментально доказал рефлекторную природу всех без исключения форм жизнедеятельности организма. По мнению И.П.Павлова, простая форма деятельности нервной системы, которая является постоянной, прирожденной, видовой и для формирования структурных предпосылок которой не требуется специальных условий, является безусловным рефлексом. Приобретаемые же в процессе жизнедеятельности временные связи, позволяющие организму устанавливать достаточно сложные и многообразные отношения с окружающей средой являются, по определению И.П.Павлова, условно рефлекторными. Местом замыкания условных рефлексов является кора больших полушарий головного мозга. Таким образом, головной мозг и его кора – основа высшей нервной деятельности.
Еще один ученый – П.К.Анохин и его ученики подтвердили наличие так называемой обратной связи рабочего органа с нервными центрами (данное явление названо «обратной афферентацией»). В тот момент, когда из центральной нервной системы эфферентные импульсы достигают исполнительных органов, в них вырабатывается ответная реакция (движение или секреция). Этот рабочий эффект раздражает рецепторы самого исполнительного органа. Возникшие в результате этих процессов импульсы по афферентным путям направляются обратно в центры спинного или головного мозга в виде информации о выполнении органом определенного действия в каждый данный момент. Таким образом, создается возможность точного учета правильности исполнения команд в виде нервных импульсов, поступающих в рабочим органам из нервных центров, и осуществляется постоянная их коррекция. Существование двусторонней сигнализации по замкнутым, круговым или кольцевым рефлекторным нервным цепочкам «обратной афферентации» позволяет производить постоянные, непрерывные, ежемоментные коррекции любых реакций организма на любые изменения условий внутренней и внешней среды. Без механизмов обратной связи приспособление живых организмов к окружающей среде было бы невозможно.
Таким образом, по мере научного прогресса на смену старым представлениям о том, что в основе деятельности нервной системы лежит «разомкнутая» (незамкнутая) рефлекторная дуга, пришло представление о замкнутой, кольцевой дуге, представляющей собой цепь рефлексов.
Процесс формирования классического условного рефлекса проходит три основные стадии.
1. Стадия прегенерализации . Характеризуется выраженной концентрацией возбуждения (главным образом в проекционных зонах коры условного и безусловного раздражителей) и отсутствием условных поведенческих реакций.
2. Стадия генерализации условного рефлекса, в основе которой лежит процесс «диффузного» распространения (иррадиации) возбуждения. Условные реакции возникают на сигнальный и другие раздражители (явление афферентной генерализации), а также в интервалах между предъявлениями условного сигнала (межсигнальные реакции). В этот период широко распространены по коре и подкорковым структурам различные биоэлектрические сдвиги (блокада альфа-ритма, десинхронизация и пр.).
3. Стадия специализации , когда межсигнальные реакции угасают и условный ответ возникает только на сигнальный раздражитель. Изменения биотоков более ограничены и приурочены в основном к действию условного стимула. Этот процесс обеспечивает дифференцировку, тонкое различение стимулов, специализацию условно-рефлекторного навыка. В процессе специализации сфере распространения биопотенциалов значительно сужена и возрастает условно-рефлекторный ответ.
Согласно результатам исследований И.П.Павлова, временная связь образуется между корковым центром безусловного рефлекса и корковым центром анализатора, на рецепторы которого действует условный раздражитель, т.е. связь замыкается в коре большого мозга). В основе замыкания временной связи лежит процесс доминантного взаимодействия между возбужденными центрами. Импульсы, вызываемые индифферентным (условным) сигналом с любого участка кожи и других органов чувств (глаз, ухо и т.д.), поступают в кору мозга и обеспечивают там образование очага возбуждения. Если после индифферентного сигнала подается пищевое подкрепление (подкормка), то возникает более мощный второй очаг возбуждения в коре больших полушарий, к которому направляется ранее возникшее и иррадиирующее по коре возбуждение. Неоднократное сочетание индифферентного (условного) сигнала и безусловного раздражителя (подкрепления) облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса.
Образование временной связи в коре больших полушарий И.П.Павлов называл замыканием новой условно-рефлекторной дуги.
Также исследованиями ученых доказано, что параллельно со становлением условного рефлекса идет процесс формирования другой условно-рефлекторной связи, специально изменяющей состояние нейронов, что выражается в росте их фоновой активности. Если по каким-либо причинам условно-рефлекторное изменение состояния данного нейрона не возникает, то выработанный у него рефлекс не обнаруживается. Это дало возможность ученым сделать вывод о том, что ассоциативный ответ включает в себя формирование состояния, качественно специфического для каждой временной связи . Данное явление рассматривается физиологами как еще один из ведущих механизмов формирования условно-рефлекторного поведения.
Таким образом, по мнению И.П.Павлова, существует два механизма условно-рефлекторной деятельности:
1. настроечный, регулирующий состояние мозга и создающий определенный уровень возбудимости и работоспособности нервных центров:
2. запускающий, который инициирует ту или иную условную реакцию.
Современное объяснение механизма образования условных рефлексов основано на представлении о модификации деятельности синапсов, существующих в тех условных пунктах нейронной сети, которые способны ассоциировать совпадавшие во времени сенсорные сигналы.
Также исследованиями ученых доказано, что процесс образования условных рефлексов непосредственно связан с памятью. В начале выработки условного рефлекса связь осуществляется только с помощью механизмов кратковременной памяти – распространение возбуждения осуществляется между двумя возбужденными корковыми центрами. По мере повторения действия условного и безусловного раздражителей и повторяющегося возбуждения соответствующих центров кратковременная память переходит в долговременную, то есть происходят значительные структурные изменения в нейронах.
Условные рефлексы, как показано многочисленными исследованиями, изменчивы (вариабельны), они могут затормаживаться.
Можно выделить два вида торможения условных рефлексов, принципиально отличающихся друг от друга: врожденное и приобретенное Рис.3). При этом каждый вид торможения имеет свои собственные вариации.
Безусловное Условное (внутреннее)
1.Внешнее 1. Угасательное
3. Дифференцировочное
4.Условный тормоз
Рис. 3. Торможение условных рефлексов
Безусловное (врожденное) торможение условных рефлексов подразделяется на внешнее и запредельное. Внешнее торможение проявляется в ослаблении или полном прекращении наличного (протекающего в данный момент) условного рефлекса при действии какого-либо постороннего раздражителя. Например, включение света во время текущего условного рефлекса вызывает появление ориентировочно-исследовательской реакции, ослабляющей или прекращающей наличную условно-рефлекторную деятельность. Эту реакцию, возникшую на изменение внешней среды (рефлекс на новизну), И.П.Павлов называл рефлексом «что такое?». С повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, поскольку организму не требуется предпринимать каких-либо действий. И.П.Павловым был изучен и механизм данного вида торможения условных рефлексов. Согласно его теории, посторонний сигнал сопровождается появлением в коре больших полушарий мозга нового очага возбуждения, который при средней силе раздражителя оказывает угнетающее воздействие на текущую условно-рефлекторную деятельность по механизму доминанты. Внешнее торможение является безусловно-рефлекторным. Внешним данный вид торможения назвали поскольку в этих случаях возбуждение клеток ориентировочно-исследовательского рефлекса, возникающего от постороннего раздражителя, находится вне дуги наличного условного рефлекса. Внешнее торможение способствует экстренному приспособлению организма к изменяющимся условиям внешней и внутренней среды и обеспечивает возможность переключиться на другую деятельность в соответствии с ситуацией.
Запредельное торможение возникает при длительном нервном возбуждении организма, при действии чрезвычайно сильного условного сигнала или нескольких несильных. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие – «закон силы»: чем сильнее условный сигнал, тем сильнее условно-рефлекторная реакция. Однако данный закон может сохраняться лишь до определенной величины (порога), выше которой эффект начинает уменьшаться, несмотря на продолжающееся увеличение силы условного сигнала. Эти факты позволили И.П.Павлову сделать вывод о том, что корковые клетки имеют предел работоспособности.
Условное (внутреннее, приобретенное) торможение условных рефлексов – это активный нервный процесс, который требует выработки, как и сам рефлекс. Не случайно данный вид торможения условного рефлекса получил название условно-рефлекторного торможения. Оно является приобретенным, индивидуальным. Согласно теории И.П.Павлова, оно локализовано в пределах («внутри») нервного центра данного условного рефлекса. Различают следующие виды условного торможения: угасательное, запаздываельное, дифференцировочное и условный тормоз.
Угасательное торможение возникает при повторном применении условного сигнала и дальнейшем его неподкреплении. При этом вначале условный рефлекс ослабевает, а затем полностью исчезает. Однако через некоторое время он может восстановиться. Скорость угасания зависит от интенсивности условного сигнала и биологической значимости подкрепления. Чем они значительнее, тем труднее совершается угасание условного рефлекса. Именно угасательным торможением можно объяснить забывание полученной ранее информации, которая длительное время не повторяется.
Запаздывательное торможение возникает при отставании подкрепления на 1-3 мин относительно начала действия условного сигнала. Постепенно появление условной реакции сдвигается к моменту подкрепления. Данному виду торможения условного рефлекса также присуще явление растормаживания.
Дифференцировочное торможение вырабатывается при дополнительном включении раздражителя, близкого к условному, и не подкреплении его.
Условный тормоз возникает при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Так, если выработать условный слюноотделительный рефлекс на свет, затем к этому сигналу подключить дополнительный раздражитель (звук), и не подкреплять эту комбинацию, то на нее условный рефлекс постепенно угаснет.
Значение всех видов условного (внутреннего) торможения условных рефлексов заключается в устранении ненужной в данное время деятельности, то есть очень тонком приспособлении организма к окружающей среде.
Зафиксированную систему условных и безусловных рефлексов, объединенных в единый функциональный комплекс, принято называть динамическим стереотипом . Динамический стереотип образуется под влиянием стереотипно повторяющихся изменений и воздействий внешней и внутренней среды организма. Повторяющиеся в одинаковой последовательности раздражители, действующие на организм, представляют собой внешний стереотип . Ему соответствует стереотипная динамика корковых процессов возбуждения и торможения, которая в результате многократных повторений внешнего стереотипа начинает воспроизводиться в одной и той же последовательности как единое целое. После этого стереотипную последовательность корковых процессов можно вызвать не только действием внешнего стереотипа (то есть комплекса раздражителей), но и действием какого-либо одного раздражителя из этого комплекса.
Понятие «динамический стереотип» было введено в начале 30-х годов ХХ столетия, когда И.П.Павлов, доказывая свою позицию относительно рефлекторной теории функционирования нервной системы. Оппонентами отечественного ученого выступали, в основном, зарубежные исследователи, которые утверждали, что рефлекторная теория перестала способствовать пониманию функций головного мозга и сделалась препятствием на пути прогресса в этой области знаний. Защищая и разъясняя свой подход к теории рефлексов, И.П.Павлов выделил в рефлекторной деятельности «три основных принципа точного научного исследования»:
· принцип детерминизма, то есть повода, причины для всякого данного действия, эффекта;
· принцип анализа и синтеза, то есть первичного разложения целого на части, составляющие единицы и затем снова постепенного сложения целого из единиц, отдельных элементов;
· принцип структурности, то есть расположения действий силы в пространстве. Данный принцип И.П.Павлов комментирует следующим образом. Когда в коре и ближайшей подкорке какой-либо раздражитель вызывает возбуждение или торможение клеток, то расположенные в разных ее частях возбужденные и тормозимые клетки образуют динамическую комбинацию друг с другом. Поскольку количество раздражителей и вариантов их комбинирования неисчислимо, то и динамические комбинации возбужденных и тормозимых клеток также не поддаются учету. Такие комбинации могут сделаться устойчивыми и существовать во время действия раздражителя. Вместе с тем, они могут сохраняться в качестве «отпечатков действительности» и после прекращения внешнего воздействия. Это означает, что след от предшествующих воздействий способен повлиять на характер ответных реакций в дальнейшем, которые, таким образом, будут зависеть не только от непосредственного стимула, но и от ранее усвоенного опыта.
И.П.Павлов рассматривал образование и сохранение динамического стереотипа как «серьезный нервный труд, различный, смотря по сложности стереотипа и индивидуальности животного».
В лаборатории И.П.Павлова использовались разнообразные схемы выработки динамических стереотипов, одни из которых были относительно просты и состояли, например, всего из двух положительных рефлексов. Другие – представляли собой сложные комбинации положительных, то есть возбуждающих, и тормозных раздражителей. Перестановка действующих раздражителей комплекса, изменение значения отдельных раздражителей с возбуждающего на тормозное или наоборот позволяли выявлять индивидуальные особенности поведения животных. В процессе изменения динамического стереотипа все животные становились повышенно возбужденными, переставали реагировать на прежние условные раздражители, иногда отказывались от пищи и сопротивлялись введению в лабораторное помещение. И.П.Павлов называл такое состояние для животного «мучительным» и объяснял его «напряженным нервным трудом», который рассматривал не только как ассоциативную деятельность, но и как деятельность (труд) умственную.
Вопросы для самоконтроля:
1. Дайте определение рефлекса.
2. Раскройте основные положения рефлекторного принципа деятельности ЦНС.
3. Какие виды рефлексов существуют?
4. Каковы специфические особенности безусловных рефлексов.
5. Раскройте механизм образования условных рефлексов.
6. Классификация условных рефлексов.
7. Механизм торможения условных рефлексов.
8. Какова роль рефлексов в жизни живых организмов?
9. Что такое рефлекторная дуга?
10. Каково строение рефлекторной дуги?
11. Охарактеризуйте простейшую рефлекторную дугу?
12. Раскройте механизм функционирования сложной рефлекторной дуги.
13. Что такое «обратная афферентация»?
14. В чем сущность и значение механизмов обратной связи?
15. Раскройте стадии формирования классического условного рефлекса.
16. Механизм торможения условных рефлексов.
17. Что такое «закон силы»?
18. Каково значение торможения условного рефлекса?
19. Что такое динамический стереотип?
Сущность работы нервной системы заключается в организации реакций в ответ на внешние и внутренние воздействия. Степень сложности таких реакций весьма различна - от автоматического сужения зрачка при ярком освещении до многопланового поведенческого акта, мобилизующего все системы организма. Тем не менее во всех случаях сохраняется один и тот же принцип деятельности - рефлекторный. Рефлекс - это активная ответная реакция, связывающая особенности организма и условия среды. Следовательно, рефлекс - не механический, не пассивный ответ, как, например, образование вмятины от удара, а целесообразная для данного организма реакция, необходимая для нормальной жизнедеятельности.
Возникновение и развитие нервной системы в процессе эволюции означало прежде всего появление и усовершенствование рефлекторных механизмов. Эти механизмы, независимо от степениих сложности, имеют ряд принципиально общих черт. Для осуществления рефлекса необходимы, как минимум, два элемента: воспринимающий (рецепторный) и исполнительный (эффекторный). Рецепторы могут реагировать на очень широкий диапазон раздражителей и занимать большие площади (рефлексогенная зона). К таким относятся, например, рецепторы болевой чувствительности, рецепторы внутренних органов. Другие воспринимающие элементы, напротив, являются чрезвычайно специализированными и имеют ограниченную рефлексогенную зону. В качестве примера можно назвать вкусовые рецепторы, располагающиеся на поверхности языка, или зрительные палочки и колбочки.
Точно так же исполнительный аппарат рефлекса может представлять собой изолированную мышцу и иметь жесткую связь с ограниченной группой рецепторов. Классический пример этого - коленный рефлекс (узкая рефлексогенная зона и элементарная реакция).В других случаях исполнительный аппарат включает в себя ансамбль действующих единиц и имеет связи с различными типами рецепторов. Примером этого может служить так называемый "стартовый" рефлекс. Он выражается в виде общего настораживания, замирания или вздрагивания при резком звуке или ярком свете, неожиданном зрительном образе. Таким образом, в реализации “стартового” рефлекса участвует огромное количество двигательных единиц и вызывается он различными раздражителями
главная особенность которых - неожиданность.
“Стартовый” рефлекс - одна из многих реакций, требующих согласованной работы различных систем организма. Такая заинтересованность невозможна при наличии жестких прямых связей с рецепторами и эффекторами, поскольку это привело бы к появлению независимых друг от друга и не поддающихся координации рефлекторных механизмов.
В процессе эволюции сформировался еще один элемент, обеспечивающий рефлекторные реакции, - вставочные нейроны. Благодаря этим нейронам импульсы от рецепторов достигают эффекторных аппаратов не сразу, а после промежуточной обработки во время которой и устанавливается согласованность в различных реакциях. Широко взаимодействуя между собой и образуя скопления, вставочные нейроны создают возможность для объединения всех рефлекторных механизмов в единое целое. Формируется интегральная нервная деятельность, которая представляет собой нечто большее, чем сумма отдельных реакций.
Каждая отдельная реакция подчиняется центральным влияниям; она может быть усилена, заторможена, полностью блокирована или приведена в состояние повышенной готовности. Более того, на основе врожденных автоматизмов формируются новые способы реагирования, новые действия. Так, ребенок обучается ходьбе, стоянию на одной ноге, сложным ручным манипуляциям.
Интегральная нервная деятельность еще не означает высшей нервной деятельности. Объединение организма в единое целое и организация сложных поведенческих программ могут совершаться на базе эволюционно закрепленных в нервной системе врожденных механизмов. Эти механизмы называются безусловными рефлексами, поскольку они генетически заложены в нервной системе и не требуют обучения. На основе безусловных рефлексов могут формироваться сложнейшие действия. В качестве примера достаточно назвать строительную деятельность бобров или дальние перелеты птиц.
Однако безусловнорефлекторная деятельность неизбежно страдает ограниченностью, потому что она почти не поддается исправлениям и тем самым препятствует накоплению индивидуального опыта. Каждый индивид от рождения почти полностью готов к определенным действиям, однообразно повторяющимся из поколения в поколение. Если условия среды внезапно изменяются. то великолепно отлаженный механизм реагирования оказывается неприспособленным.
Гораздо большая гибкость поведения наблюдается у организмов, которые способны к индивидуальному обучению. Это становится возможным благодаря возникновению в нервной системе временных нервных связей. Наиболее изученным типом такой нервной связи является условный рефлекс. При помощи этого рефлекса раздражитель, бывший ранее безразличным, приобретает значение жизненно важного сигнала и вызывает определенную реакцию. В механизмах условного рефлекса заложены предпосылки индивидуальной памяти, без которой, как известно, невозможно обучение.
По мере эволюционирования коры больших полушарий возникают огромные зоны нервных клеток, которые не имеют никакой врожденной программы, а предназначены лишь для образования связей в процессе индивидуального обучения. Поскольку работа нервной системы основана на рефлекторном принципе, то и обучение распространяется на три основные звена рефлекторного механизма: анализ поступающей от рецепторов информации, интегральная обработка в промежуточных звеньях, создание новых программ деятельности.
Личный опыт оказывает влияние как на восприятие и переработку информации из внешней и внутренней среды, так и на формирование программ деятельности - краткосрочных или долгосрочных. В результате восприятия многих раздражителей происходит опознавание, т.е. сведения о раздражителе сравниваются с заложенной в памяти информацией. Точно так же при организации ответных действий учитываются не только потребности на данный момент, но и прошлый опыт успешных или неуспешных реакций в аналогичной ситуации.
При выполнении намеченного действия могут возникнуть непредвиденные помехи. Следовательно, необходимо сохранять конечную цель реакции до ее полного осуществления, для чего требуются специальные механизмы.
Процессы распознавания поступающих сигналов, выработка учитывающих прошлый опыт программ действия, контроль за их выполнением составляют содержание высшей нервной деятельности. Эта деятельность, оставаясь рефлекторной по своей сущности, отличается от врожденных автоматизмов гораздо большей гибкостью и избирательностью. Один и тот же раздражитель может вызывать разные реакции в зависимости от состояния на данный момент, общей ситуации, индивидуального опыта, потому что многое зависит не от особенностей раздражителя, а от той обработки, которую он проходит в промежуточных звеньях рефлекторного аппарата.
Высшая нервная деятельность создает предпосылки разума. Разум означает прежде всего способность найти решение в новой необычной ситуации. Приведем пример. Обезьяна видит подвешенную к потолку связку бананов и разбросанные по полу ящики. Без предварительного обучения она решает возникшую перед ней практическую и интеллектуальную задачу - ставит один ящик на другой и достает бананы. С возникновением речи возможности интеллекта безгранично расширяются, поскольку в словах отражена сущность окружающих нас вещей.
Высшая нервная деятельность является нейрофизиологической основой психических процессов. Но она их не исчерпывает. Для таких психических явлений, как чувство, воля, воображение, мышление, конечно, необходима соответствующая мозговая активность Однако конкретное содержание психических процессов определяется социальной средой, а не процессами возбуждения или торможения в нейронах. Решает ли ученый сложнейшую интеллектуальную задачу или же первоклассник обдумывает простенькую школьную задачку, их мозговая активность может быть примерно одинаковой. Направленность мозговой деятельности задается не физиологией нервных клеток, а смыслом выполняемой работы.
Однако сказанное не означает, что высшая нервная деятельность представляет собой нечто второстепенное по отношению к “истинно психическим” процессам. Напротив, общие закономерности взаимодействия нейронов и общие принципы организации нервных центров определяют многие характеристики психической деятельности, например, темпы интеллектуальной работы, устойчивость внимания, объем памяти. Эти и другие показатели имеют огромное значение для педагогической работы, особенно при наличии у детей дефектов центральной нервной системы.
Сложнейшие мозговые механизмы, обеспечивающие переработку информации, поступающей сразу от многих рецепторных зон и промежуточных центров, представляют большой интерес как для физиологии, так и для психологии. Наблюдается все большее взаимопроникновение этих двух дисциплин, что отражается и на учении о высшей нервной деятельности.
В учении о высшей нервной деятельности можно выделить два основных раздела. Первый из них стоит ближе к нейрофизиологии и рассматривает общие закономерности взаимодействия нервных центров, динамику процессов возбуждения и торможения. Второй раздел рассматривает конкретные механизмы отдельных мозговых функций, таких как речь, память, восприятие, произвольные движения, эмоции. Этот раздел близко примыкает к психологии и нередко обозначается как психофизиология. Кроме того, произошло выделение самостоятельного направления - нейропсихологии. Нейропсихология в значительной степени - клиническая дисциплина. Она не только изучает механизмы высших корковых функций, но и разрабатывает методы точной диагностики корковых поражений и принципы коррекционных мероприятий. Один из основателей нейропсихологии - выдающийся отечественный ученый А. Р. Лурия.
Названные разделы тесно взаимосвязаны, поскольку мозг работает как единое целое. Однако для наилучшего понимания общих закономерностей высшей нервной деятельности целесообразно рассмотреть по отдельности принципы высшей нейродинамики и нейропсихологические механизмы отдельных корковых функций.
ДИНАМИКА НЕРВНЫХ ПРОЦЕССОВ
Принципы высшей нейродинамики - это закономерности взаимодействия процессов возбуждения и торможения в клетках головного мозга. Основные закономерности таких процессов были раскрыты И.П.Павловым и его учениками.
Возбуждение и торможение способны иррадиировать, т.е. распространяться на новые клеточные зоны, и концентрироваться, т.е. ограничиваться определенным очагом. Процессы иррадиации иконцентрации обусловливают пеструю и постоянно меняющуюся мозаику распределения возбужденных и заторможенных мозговых участков.
Степень иррадиации возбуждения зависит от многих факторов: силы раздражителя, его новизны, значимости для организма. Кроме того, большое значение имеет закон отрицательной индукции - возникновение зоны торможения вокруг очага возбуждения. Отрицательная индукция препятствует безграничной иррадиации возбуждения. В противном случае каждый раздражитель полностью “захватывал” бы огромные массы клеток. Такая картина наблюдается при судорожном припадке: очаг возбуждения безудержно распространяется на все новые и новые зоны; сознание при этом обычно утрачивается.
Иррадиация и концентрация возбуждения лежат в основе механизма внимания. Объем и стойкость внимания зависят от величины очага возбуждения и его фиксированности. Способность произвольно контролировать направленность, объем и устойчивость внимания совершенствуется с возрастом. Внимание детей характеризуется слабой целенаправленностью, но большим объемом. Дети фиксируют множество деталей автоматически; взрослые направляют внимание более прицельно, но и более узко. Кроме того, внимание детей неустойчиво. Это обусловлено недостаточным Развитием внутреннего торможения, обеспечивающего дополнительную концентрацию внимания. Каждый новый раздражитель отвлекает ребенка. Здесь опять-таки проявляется принцип отрицательной индукции: новый очаг возбуждения тормозит сущестовавший до этого. У взрослых процессы возбуждения и торможения более сбалансированны, поэтому возникающие конкурентные очаги возбуждения блокируются. Это достигается прежде всего за счет взаимодействия лобных долей мозга и ретикулярной формации. При поражениях лобных долей наблюдается чрезмерная отвлекаемость больных: их внимание постоянно переключается с одного объекта на другой.
Наряду с отрицательной индукцией существует положительная - возникновение возбуждения вокруг очага торможения. Например, засыпающий человек, многие участки мозга которого заторможены, вдруг начинает отчетливо слышать тиканье часов шум капающей из крана воды и другие звуки, не замечавшие в бодрствующем состоянии. Вероятно, это объясняется возникновением активных очагов на фоне общего снижения бодрствования.
В головном мозге обычно одновременно существует значительное количество возбужденных очагов. При этом может возникнут ситуация, когда какой-то один очаг начинает не только подавлять все остальные, но и использовать их активность для усиления своей собственной. Формируется так называемая доминанта, которую подробно изучил выдающийся отечественный физиолог А. А. Ухтомский. Доминанта - это очаг активности, подчиняющий себе все остальные, даже не имеющие к нему непосредственного отношения. Например, изголодавшемуся человеку все напоминает о еде, даже, казалось бы, совершенно посторонние разговоры и предметы. Точно так же увлеченный какой-либо идеей ученый находит тему для размышлений в событиях и фактах, относящихся к весьма отдаленным областям.
Принцип доминанты имеет важное биологическое значение, позволяя организму достигать необычайной концентрации усилий для выполнения какой-либо жизненно необходимой задачи Благодаря доминанте различные отвлекающие факторы не мешают, а, напротив, усиливают стремление к главной цели. Однако доминанта может принимать и патологические черты, если она направлена на утратившие значение или вообще не имеющие смысла цели. Такая картина, в частности, наблюдается при бредовых идеях. Больной не только уверен в правильности своих нелепых мыслей, но в ответ на возражения еще больше убеждается в своей правоте. Переубедить человека с бредовой идеей практически невозможно.
По мере достижения поставленной цели физиологическая доминанта обычно угасает. У человека длительное постоянство устремлений поддерживается благодаря усилиям воли.
Как уже отмечалось, степень иррадиации процессов возбуждения и торможения зависит не только от интенсивности раздражителей, но и от их значимости. Эта значимость может быть безусловно-рефлекторной, основанной на врожденной способности реагирования, но может обусловливаться и индивидуальным опытом Например, собака поразному реагирует на черствый кусок хлеба и на аппетитную кость. Это врожденная способность оценивать качество пищи. В то же время в процессе обучения любая собака приобретает большой опыт по распознаванию “пищевой ценности" различных раздражителей (хлопанье дверцы холодильника, звяканье посуды и др.). Процесс превращения безразличного ранее раздражителя в значимый для организма сигнал блестяще изучен И.П. Павловым. В многочисленных экспериментах И. П. Павлов и его ученики показали, что если перед безусловнорефлекторным раздражителем предъявлять какой-либо другой, то после ряда повторений этот раздражитель способен самостоятельно вызывать данную безусловную реакцию. Вырабатывается так называемый условный рефлекс, вызывающийся раздражителем, который до эксперимента был безразличен для животного. Открытие условного рефлекса показало, каким образом индивидуальный опыт фиксируется в виде нервных связей, как происходит элементарное обучение. Было установлено, что в процессе образования условных рефлексов большую роль играют процессы торможения. В частности, большое значение имеет так называемое дифференцировочное торможение, благодаря которому характеристики условнорефлекторного раздражителя оцениваются более точно. Например, при выработке условной слюноотделительной реакции на звук звонка первоначально реакция возникает в ответ на любой звонок. В дальнейшем, если подкреплять кормление только звонком определенной тональности и продолжительности, слюноотделительный рефлекс становится более избирательным: уже далеко не всякий звук вызывает слюноотделение. Этот факт свидетельствует о наличии выборочного торможения сходных сигналов в зависимости от прошлого опыта.
Дифференцировочное торможение И. П. Павлов относил к разновидностям внутреннего. Его существование указывает на способность к значительному усовершенствованию реагирования.
Существование внутреннего торможения обнаруживается также при выработке так называемых отставленных условных рефлексов. Суть их заключается в том, что после предъявления условного раздражителя подкрепление дается не сразу, а спустя некоторое время. В результате, например, слюноотделение в ответ на звонок возникает не сразу, а через некоторый промежуток времени. В течение всего периода между предъявлением звонка и появлением слюны реакция тормозится.
Внутреннее торможение играет большую роль в процессе обучения, совершенствования поведения. В известной степени воспитание сводится к тренировке внутреннего торможения, поскольку именно оно обеспечивает гибкость и тонкость реакций.
Внутреннее торможение требует больших усилий от нервной системы. В экспериментах на животных неоднократно наблюдалось, как при выработке слишком тонкой дифференцировки (например, между кругом и почти круглым овалом) или при чрезмерном временном разрыве между сигналом и подкреплением животное сильно возбуждалось, начинало вырываться из станка, проявляло агрессивность. В других случаях, наоборот, наступало оцепенение, возникала непреодолимая сонливость. Кстати, сонливость здесь является результатом так называемого запредельного торможении, которое распространяется по нервной системе при непосильных нагрузках и предохраняет нервные клетки от истощения.
Приведенные примеры говорят о том, что тренировка внутреннего торможения требует строго дозированных нагрузок. В противном случае может произойти срыв, дезорганизация высшей нервной деятельности. Подобные явления иногда наблюдаются в школе при изложении чрезмерно сложного материала. Одни ученики становятся невнимательными, непоседливыми, начинают разговаривать. Других же клонит в сон, они зевают, усиленно моргают. При наличии дефектов центральной нервной системы способность к выработке внутреннего торможения ограниченна, что делает необходимым более тщательное дозирование учебных нагрузок.
В процессе изучения условных рефлексов было установлено что они могут приобретать тормозное значение, блокировать отдельные реакции или вызывать сон. Таким образом было открыто условное торможение, которое И. П. Павлов относил к разновидностям внешнего торможения, поскольку оно вызывается сигналом из внешней среды. Условное торможение имеет важное значение в регуляции ритма сна - бодрствования. Систематически повторяющаяся процедура приготовления ко сну представляет собой, по существу, набор условных рефлексов, облегчающих засыпание. В организации режима дня ребенка важно добиваться строгого повторения такой процедуры, поскольку многие дети укладываются спать очень неохотно.
Другой разновидностью внешнего торможения является запредельное, о котором уже говорилось. Однако запредельное торможение по своей природе безусловнорефлекторное, оно представляет собой врожденное свойство нервной системы. В животном мире широко распространена так называемая реакция “мнимой смерти” - в случае опасности животное замирает, становится словно парализованным. У людей подобные реакции обозначаются как реактивный ступор, который может продолжаться и спустя несколько суток после потрясения. Частным случаем такого ступора является реактивный мутизм - утрата речи при сохранении общей двигательной способности. Реактивный мутизм иногда возникает у робких, застенчивых детей, впервые выступающих перед большим скоплением незнакомых людей.
По данным многочисленных экспериментов, условные рефлексы не обязательно являются изолированной реакцией на отдельные раздражители (звонок - выделение слюны и др.). У многих животных успешно вырабатываются условные рефлексы на сложные комплексы раздражителей, воздействующих одновременно последовательно на многие рецепторные аппараты (например, свет, звук, прикосновение, запах). Кроме того, рефлекторный ответ может представлять собой набор реакций, совершающихся одновременно или разворачивающихся во времени в определенной последовательности. Например, дрессированной собаке достаточно одной команды, чтобы она совершила серию действий, сменяющихся в заданном порядке. Каждый человек в процессе воспитания и обучения усваивает массу двигательных навыков, предназначенных для выполнения обыденных житейских операций: одевание, умывание, причесывание, еда при помощи ложки и вилки, склеивание бумаги, зажигание спичек и т.д. Любое из этих действий представляет собой слитую воедино последовательность движений. Например, чтобы съесть ложку супа, необходимо придать ложке определенное положение в руке, зачерпнуть суп, донести его, не пролив, до рта и, наконец, перелить содержимое в рот. Всему этому человек учится в детстве, “отрабатывая” каждый элемент действия по отдельности: как правильно держать ложку и перемещать ее в пространстве, какое положение придать губам, чтобы ничего не пролилось. В результате формируется цепочка движений, слитых в единый автоматизированный акт, и в дальнейшем человек уже совершенно не будет задумываться о том, как пользоваться ложкой.
Прочно зафиксированная в нервной системе последовательность реакций называется динамическим стереотипом. Способность к формированию динамических стереотипов приводит к огромной экономии в работе нервной системы. Много раз повторяющиеся операции закрепляются как целостные двигательные образы, поэтому отпадает необходимость каждый раз находить способы реализации того или иного действия. Достаточно “начальнику” дать команду, и весь комплекс движений “проигрывается”, как записанная на пластинке мелодия.
Динамические стереотипы могут формироваться не только в сфере движений, но и в сфере восприятия. Например, горожанин, переходя улицу, автоматически обращает внимание на сигнал светофора, поворачивает голову налево, потом направо. На основе динамических стереотипов вырабатываются профессиональные навыки: работа с инструментами, печатание на пишущей машинке, укладка кирпича и т.д. Следует отметить, что в динамическом стереотипе могут содержаться бесполезные и даже мешающие выполнению задачи элементы. Это зависит от особенностей процесса обучения. Например, походка человека представляет собой классический динамическийий стереотип, и здесь немало отрицательных характеристик (шарканье ногами, сгорбленность, раскачивание и др.). Все что является результатом того, что в раннем детстве родители ребенка не обращали должного внимания на его походку. Между тем данные элементы стереотипа фиксируются очень прочно, искоренить их весьма трудно. При выработке нового стереотипа важно с самого начала следить за качеством его отдельных элементов. В частности, из логопедической практики хорошо известно, что дислалия часто является следствием фиксации физиологической дислалии дошкольников. Сложившийся аномальный стереотип звукопроизношения переделывают уже с помощью логопеда.
Сложнейшая динамика взаимодействия процессов возбуждения и торможения создает постоянно меняющуюся картину мозговой активности. Однако в этой переменчивости существуют некоторые стабильные характеристики, определяющие индивиду. альные особенности реагирования.
С древнейших времен известно, что одни люди отвечают на все происходящее бурными реакциями, другие же, наоборот, всегда остаются чрезвычайно спокойными. Важно подчеркнуть, что такой стиль реагирования может оставаться устойчивой характеристикой в течение всей жизни человека и, следовательно, он является врожденной особенностью.
Общий тип реагирования, определяющий стиль поведения, издавна обозначается как темперамент. Существует много классификаций темпераментов, но наибольшую известность имеет типология, описанная еще в античную эпоху.
Античная классификация темпераментов основывалась на наивном представлении о пропорциях различных жидкостей в организме. Отсюда произошли и названия четырех основных типов: холерик (холе - желчь), сангвиник (сангвис - кровь), флегматик (флегма - слизь) и меланхолик (мелан холе - черная желчь). Однако описательные характеристики этих темпераментов точно подмечали реально существующие особенности людских характеров.
Холерик - человек взрывчатый, бурно на все реагирующий, но быстро “остывающий”, легко меняющий интересы и увлечения, сангвиник - энергичный, активный, способный доводить начатое дело до конца, флегматик - спокойный, невозмутимый, медленно “раскачивающийся”, но стойкий в своих переживаниях, меланхолик - робкий, нерешительный, легко ранимый, однако способный к очень тонким переживаниям и наблюдениям.
И.П.Павловым была раскрыта нейрофизиологическая основа темпераментов. В качестве ведущих характеристик высшей нервной деятельности рассматривались сила, подвижность и уравновешенность процессов возбуждения и торможения. В зависимости от сочетания этих особенностей выделены четыре основных типа высшей нервной деятельности.
Сильный, подвижный, неуравновешенный соответствует холерическому темпераменту; сильный, подвижный, уравновешенный - сангвиническому; сильный, инертный - флегматическому слабый, тормозимый тип - меланхолическому.
Кроме того, основываясь на особенностях взаимодействия первойи второй сигнальной систем (чувственно-конкретное и речевое восприятия), И.П.Павлов выделил художественный (первосигнальный), мыслительный (второсигнальный) и средний, промежуточный типы.
Тип высшей нервной деятельности во многом определяется
врожденными свойствами нервной системы, но не является совершенно незыблемым, не поддающимся изменениям. Можно даже сказать, что почти любой ребенок в процессе развития совершает эволюцию от холерического, художественного темперамента к уравновешенному, мыслительному. Тем не менее существуют дети явно возбудимые и явно заторможенные, энергичные и пассивные, самоуверенные и робкие, выносливые и утомляемые. В связи с этим в педагогической работе важно учитывать индивидуальные особенности высшей нервной деятельности, проводя в то же время коррекцию мешающих работе характеристик. Особое значение такой подход приобретает в дефектологии, где многие дети нуждаются в специальной помощи при формировании каркаса высшей нервной деятельности.
ВЫСШИЕ КОРКОВЫЕ ФУНКЦИИ
Кора головного мозга является, по существу, гигантским промежуточным центром на пути от рецепторных аппаратов к эффекторным. Сюда стекается вся информация, поступающая из внешней и внутренней среды, здесь она сопоставляется с текущими потребностями, прошлым опытом и преобразуется в команды, нередко охватывающие все процессы жизнедеятельности. Здесь вырабатываются принципиально новые решения, а также формируются динамические стереотипы, образующие шаблоны поведения, восприятия и, в ряде случаев, даже мышления.
Связь коры с “периферическими” образованиями - рецепторами и эффекторами - обусловливает специализацию отдельных ее участков. Различные области коры связаны со строго определенными типами рецепторов, образуя корковые отделы анализаторов.
Анализатор - специализированная физиологическая система, обеспечивающая прием и переработку определенного типа раздражений. В нем различают периферический отдел - собственно pецепторные образования - и совокупность промежуточных центров. Наиболее важные центры расположены в зрительном бугре, являющемся коллектором всех видов чувствительности, и в коре больших полушарий. Корковые отделы анализаторов представляют собой высшие, но не конечные, центры, поскольку поступающие сюда импульсы не “оседают” здесь, как в хранилище, а постоянно перерабатываются, преобразуясь в командные сигналы. Эти команды могут направляться к рецепторным аппаратам, изменяя порог их чувствительности. В результате каждый анализатор функционирует как кольцевая структура, в которой импульсы циркулируют по маршруту рецепторы - промежуточные центры - рецепторы. Разумеется, что от промежуточных центров имеются выходы и к эффекторным аппаратам. Действие же эффекторов, в свой очередь, порождает новые рецепторные сигналы. В итоге формируются сложные кольцевые системы: рецептор - промежуточный центры - эффектор - рецептор. Такие системы могут иметь не сколько уровней замыкания (продолговатый, межуточный мозг, но высшим является корковый. Низшие уровни регуляции xapaктеризуются жестким автоматизмом, высшие, особенно корковые отличаются большей гибкостью и изменчивостью.
Основные корковые отделы анализаторов имеют следующее расположение (см. рис. 9): зрительный анализатор - в затылочной коре, слуховой - в височной, поверхностная и глубокая чувствительность - в задней центральной извилине, двигательный анализатор - в передней центральной извилине. Обонятельный анализатор располагается в эволюционно более древних отделах коры. включающих аммонов рог и поясную извилину. Вкусовая чувствительность и рецепция от внутренних органов имеют менее определенное корковое представительство, концентрируясь в основном в глубинных отделах сильвиевой борозды.
Каждый анализатор представлен в симметричных отделах правого и левого полушарий мозга. Двигательный и чувствительны!: анализаторы связаны с противоположной половиной тела. Корковые представительства слухового, вкусового и обонятельного анализаторов в каждом полушарии имеют связи с обеими сторонами. В зрительную кору (затылочная область) проецируется информация от половины поля зрения каждого глаза, причем в левое полушарие - от правых половин, в правое - от левых половин полей зрения.
Из анатомических особенностей следует, что расстройства движений, чувствительности и зрения возможны при поражении соответствующего участка одного из полушарий. Данные нарушения возникают на стороне, противоположной локализации патологического очага. Корковые расстройства слуха, вкуса и обоняния наблюдаются только при двустороннем поражении анализаторных зон или их связей.
Наличие симметричных анализаторных отделов в правом и левом полушариях не означает их полной равноценности. Многочисленными экспериментами доказано существование функциональной асимметрии мозга. Ее суть заключается в том, что правое и левое полушария выполняют несколько различные функции. Различают доминантное и субдоминантное полушария. В доминантном располагаются центры речи и письма, в субдоминантном соответствующие центры отсутствуют. Чаще всего доминантным полушарием является левое, и расположение в нем речевых центров обычно совпадает с праворукостью - преобладанием правой руки над левой.
В случаях выраженной леворукости доминантным может быть правое полушарие. Однако вопрос о левшестве далеко не прост.
В процессе воспитания большинство родителей приучают детей пользоваться преимущественно правой рукой. Трудно сказать, какое полушарие доминирует у “переученных левшей”. Кроме того, встречаются случаи амбидекстрии - примерно одинакового владенния обеими руками. Сложно оценивать также степень функциональной асимметрии мозга. Тем не менее эта асимметрия существует, о чем убедительно свидетельствуют результаты исследований по изолированному выключению активности правого или левого полушария, а также клинический анализ право- и левополушарных поражений мозга. Роль каждого из полушарий освещена подробнее при описании отдельных высших корковых функций.
Изучение микроскопической структуры корковых отделов анализаторов показало, что в каждом таком отделе существуют два типа клеточных зон. В центре коркового представительства анализатора располагаются первичные клеточные поля, называемые также проекционными. Их особенность состоит в том, что они имеют непосредственную связь с периферическими отделами анализатора и являются, таким образом, первыми получателями информации (или отправителями - в случае двигательного анализатора). Первичные клеточные поля отличаются высокой специфичностью, т.е. настроены на прием информации от определенных типов рецепторов. Кроме того, в этих полях нередко наблюдается и вполне определенное расположение представительств отдельных рецепторных зон. Так, в задней центральной извилине каждая часть тела имеет свою область проекции: в верхних отделах - нижняя конечность, в средних - рука, в нижних - лицо. Аналогичная картина наблюдается и в передней извилине. В зрительной коре различные квадранты полей зрения (квадрант - четвертая часть) проецируются в строго определенные участки. Таким образом, в первичных, или проекционных, зонах наблюдается высокая избирательность в приеме информации и специальная представленность отдельных рецепторных зон. В периферических отделах корковых представительств анализаторов располагаются вторичные, или проекционно-ассоциационные, клеточные зоны. Для них характерны гораздо меньшая специализированность в приеме информации и отсутствие прямой связи с периферией. В то же время эти зоны способны устанавливать контакты с другими отделами коры, а также образовывать внутри себя сложные комплексы, в которых, как считается, фиксируется прошлый опыт.
Таким образом, вторичные клеточные зоны, надстраиваясь над первичными, обеспечивают более сложную переработку информации и формируют при каждом анализаторе специализированные блоки памяти.
При оценке площади, занимаемой первичными и вторичными клеточными зонами анализаторов, нетрудно увидеть, что значительные пространства поверхности коры остаются как бы “не занятыми”. К таким “свободным” территориям относятся прежде всего обширная теменно-височно-затылочная область и участка лобной доли кпереди от передней центральной извилины. Между тем именно эти отделы коры неуклонно увеличиваются по мере эволюционирования и достигают наибольшего развития у человека. Специальные исследования показывают, что в этих отделах paсполагаются третичные корковые зоны.
Для третичных клеточных зон характерна способность к восприятию многоплановой информации; здесь отсутствует узкая специализированность. В третичных зонах осуществляется межанализаторный анализ и синтез информации, что обеспечивает комплексную память, организацию работы мозга в целом. При этом многомерный, многоплановый анализ окружающей действительности осуществляется преимущественно в височно-теменно-затылочной области, а планирование действий, разработка сложных программ поведения производится главным образом в лобной доле. Именно в третичных зонах формируются центр речи письма, счета, зрительно-пространственной ориентировки. Здесь фиксируются также навыки, приобретенные человеком в процессе его социального обучения. Важно отметить, что функционалная асимметрия мозга особенно наглядно выступает в работе третичных зон. Доминантное и субдоминантное полушария вносят неоднозначный вклад в осуществление “третично организованных" корковых функций.
Учитывая наличие различных клеточных зон, можно считать, что в коре головного мозга происходят две основные группы процессов внутриа
Основным и специфическим проявлением деятельности нервной системы является рефлекторный принцип. Это способность организма реагировать на внешние или внутренние раздражения двигательной, или секреторной реакцией. Основы учения о рефлекторной деятельности организма были заложены французским ученым Рене Декартом (1596-1650). Наибольшее значение имели его представления о рефлекторном механизме взаимоотношений организма с окружающей средой. Сам термин «рефлекс» был введен значительно позднее - в основном после выхода работ выдающегося чешского анатома и физиолога Г. Прохаски (1749-1820).
Рефлекс - это закономерная реакция организма в ответ на раздражение рецепторов, которая осуществляется рефлекторной дугой при участии центральной нервной системы. Это приспособительная реакция организма в ответ на изменение внутренней или окружающей среды. Рефлекторные реакции обеспечивают целостность организма и постоянство его внутренней среды, рефлекторная дуга является основной единицей интегративной рефлекторной активности.
Значительный вклад в развитие рефлекторной теории внес И.М. Сеченов (1829-1905). Он первым использовал рефлекторный принцип для изучения физиологических механизмов психических процессов. В работе «Рефлексы головного мозга» (1863) И.М. Сеченов аргументировано доказал, что психическая деятельность человека и животных осуществляется по механизму рефлекторных реакций, которые происходят в головном мозге, включая самые сложные из них - формирование поведения и мышление. На основании проведенных исследований он сделал вывод, что все акты сознательной и бессознательной жизни являются рефлекторными. Рефлекторная теория И.М. Сеченова послужила основой, на которой возникло учение И.П. Павлова (1849-1936) о высшей нервной деятельности. Разработанный им метод условных рефлексов расширил научное понимание роли коры большого мозга как материального субстрата психики. И.П. Павлов сформулировал рефлекторную теорию работы головного мозга, которая основывается на трех принципах: причинности, структурности, единстве анализа и синтеза. П. К. Анохин (1898-1974) доказал значение обратной связи в рефлекторной деятельности организма. Суть ее состоит в том, что во время осуществления любого рефлекторного акта процесс не ограничивается лишь эффектором, а сопровождается возбуждением рецепторов рабочего органа, от которых информация о последствиях действия поступает афферентными путями к центральной нервной системе. Появились представления о «рефлекторном кольце», «обратной связи».
Рефлекторные механизмы играют существенную роль в поведении живых организмов, обеспечивая адекватное их реагирование на сигналы окружающей среды. Для животных действительность сигнализируется почти исключительно раздражениями. Это первая сигнальная система действительности, общая для человека и животных. И.П. Павлов доказал, что для человека, в отличие от животных, объектом отображения является не только окружающая среда, но и общественные факторы. Поэтому для него решающее значение приобретает вторая сигнальная система - слово как сигнал первых сигналов.
Условный рефлекс лежит в основе высшей нервной деятельности человека и животных. Он всегда включается как существенный компонент в самых сложных проявлениях поведения. Однако не все формы поведения живого организма можно объяснить с точки зрения рефлекторной теории, которая раскрывает лишь механизмы действия. Рефлекторный принцип не дает ответа на вопрос о целесообразности поведения человека и животных, не учитывает результата действия.
Поэтому на протяжении последних десятилетий на основании рефлекторных представлений сформировалось понятие относительно ведущей роли потребностей как движущей силы поведения человека и животных. Наличие потребностей является необходимой предпосылкой любой деятельности. Деятельность организма приобретает определенную направленность лишь при наличии цели, которая отвечает данной потребности. Каждому поведенческому акту предшествуют потребности, которые возникли в процессе филогенетического развития под влиянием условий окружающей среды. Именно поэтому поведение живого организма определяется не столько реакцией на внешние воздействия, сколько необходимостью реализации намеченной программы, плана, направленных на удовлетворение той или иной потребности человека или животного.
П.К. Анохин (1955) разработал теорию функциональных систем, которая предусматривает системный подход к изучению механизмов работы головного мозга, в частности, разработки проблем структурно-функциональной основы поведения, физиологии мотиваций и эмоций. Суть концепции - мозг может не только адекватно реагировать на внешние раздражения, но и предусматривать будущее, активно строить планы своего поведения и реализовывать их. Теория функциональных систем не исключает метода условных рефлексов из сферы высшей нервной деятельности и не заменяет его чем-то другим. Она дает возможность глубже вникать в физиологическую сущность рефлекса. Вместо физиологии отдельных органов или структур мозга системный подход рассматривает деятельность организма в целом. Для любого поведенческого акта человека или животного нужна такая организация всех мозговых структур, которая обеспечит нужный конечный результат. Итак, в теории функциональных систем центральное место занимает полезный результат действия. Собственно факторы, которые находятся в основе достижения цели, формируются по типу разносторонних рефлекторных процессов.
Одним из важных механизмов деятельности центральной нервной системы является принцип интеграции. Благодаря интегрированию соматических и вегетативных функций, которое осуществляется корой большого мозга через структуры лимбико-ретикулярного комплекса, реализуются разнообразные приспособительные реакции и поведенческие акты. Высшим уровнем интеграции функций у человека являются лобные отделы коры.
Важную роль в психической деятельности человека и животных играет принцип доминанты, разработанный О. О. Ухтомским (1875-1942). Доминанта (от лат. dominari господствовать) это превосходящее в центральной нервной системе возбуждение, которое формируется под влиянием стимулов окружающей или внутренней среды и в определенный момент подчиняет себе деятельность других центров.
Головной мозг с его высшим отделом - корой большого мозга - это сложная саморегулировочная система, построенная на взаимодействии возбудительных и тормозных процессов. Принцип саморегуляции осуществляется на разных уровнях анализаторных систем - от корковых отделов до уровня рецепторов с постоянным подчинением низших отделов нервной системы высшим.
Изучая принципы функционирования нервной системы, не без основания головной мозг сравнивают с электронной вычислительной машиной. Как известно, основой работы кибернетического оснащения являются прием, передача, переработка и сохранение информации (память) с дальнейшим ее воспроизведением. Для передачи информация должна быть закодирована, а для воспроизведения - раскодирована. Пользуясь кибернетическими понятиями, можно считать, что анализатор принимает, передает, перерабатывает и, возможно, сохраняет информацию. В корковых отделах осуществляется ее раскодирование. Это, наверное, достаточно, чтобы сделать возможной попытку сравнить мозг с компьютером. Вместе с тем нельзя отождествлять работу головного мозга с вычислительной машиной: «...мозг - наиболее капризная машина в мире. Будем же скромными и осторожными с выводами» (И.М. Сеченов, 1863). Компьютер - это машина и ничего больше. Все кибернетические устройства работают по принципу электрического или электронного взаимодействия, а в головном мозге, который создан путем эволюционного развития, кроме того, происходят сложные биохимические и биоэлектрические процессы. Они могут осуществляться только в живой ткани. Головной мозг, в отличие от электронных систем, функционирует не по принципу «все или ничего», а учитывает великое множество градаций между этими двумя крайностями. Эти градации обусловлены не электронными, а биохимическими процессами. В этом существенное отличие физического от биологического. Головной мозг имеет качества, которые выходят за пределы тех, которые имеет вычислительная машина. Следует добавить, что поведенческие реакции организма в значительной мере определяются межклеточным взаимодействием в центральной нервной системе. К одному нейрону, как правило, подходят отростки от сотен или тысяч других нейронов, и он, в свою очередь, ответвляется в сотни или тысячи других нейронов. Никто не может сказать, сколько в мозге синапсов, но число 10 14 (сто триллионов) не кажется невероятным (Д. Хьюбел, 1982). Компьютер вмещает значительно меньше элементов. Функционирование головного мозга и жизнедеятельность организма осуществляются в конкретных условиях окружающей среды. Поэтому удовлетворение тех или иных потребностей может быть достигнуто при условии адекватности этой деятельности существующим внешнесредовым условиям.
Для удобства изучения основных закономерностей функционирования головной мозг разделяют на три основные блока, каждый из которых выполняет свои определенные функции.
Первый блок - это филогенетически древнейшие структуры лимбико-ретикулярного комплекса, которые расположены в стволовых и глубинных отделах головного мозга. В их состав входят поясная извилина, морской конек (гиппокамп), сосочкоподобное тело, передние ядра таламуса, гипоталамус, сетчатая формация. Они обеспечивают регуляцию жизненно необходимых функций - дыхания, кровообращения, обмена веществ, а также общего тонуса. Относительно поведенческих актов, то эти образования принимают участие в регуляции функций, направленных на обеспечение пищевого и сексуального поведения, процессов сохранения вида, в регуляции систем, которые обеспечивают сон и бодрствование, эмоциональную деятельность, процессы памяти.
Второй блок - это совокупность образований, размещенных позади центральной борозды: соматосенсорные, зрительные и слуховые зоны коры большого мозга. Основные их функции: прием, переработка и сохранение информации.
Нейроны системы, которые размещены преимущественно кпереди от центральной борозды и связаны с эффекторными функциями, реализацией двигательных программ, составляют третий блок.
Тем не менее следует признать, что нельзя провести четкой границы между сенсорными и моторными структурами мозга. Постцентральная извилина, которая является чувствительной проекционной зоной, тесно взаимосвязана с прецентральной двигательной зоной, образовывая единое сенсомоторное поле. Поэтому необходимо четко понимать, что та или другая деятельность человека требует одновременного участия всех отделов нервной системы. Причем система в целом выполняет функции, которые выходят за пределы функций, присущих каждому из указанных блоков.
Ответная реакция организма на раздражение из внешней или внутренней среды, осуществляющаяся при участии ЦНС, называется рефлексом. Путь, по которому проходит нервный импульс от рецептора до эффектора (действующий орган), называется рефлекторной дугой.
В рефлекторной дуге (рис. 104) различают пять звеньев: 1) рецептор; 2) чувствительное волокно, проводящее возбуждение к центрам; 3) нервный центр, где происходит переключение возбуждения с чувствительных клеток на двигательные; 4) двигательное волокно, передающее нервные импульсы на периферию; 5) действующий орган - мышца или железа.
Рис. 104. Схема рефлекторной дуги. А - соматического рефлекса; Б -
вегетативного рефлекса; 1 - рецептор; 2 - чувствительный нейрон; 3 -
центральная нервная система; 4 - двигательный нейрон; 5 - рабочий
орган - мышца, железа; 6 - ассоциативный (вставочный) нейрон; 7 -
вегетативный узел (ганглий)
Любое раздражение: механическое, световое, звуковое, химическое, температурное, воспринимаемое рецептором, трансформируется (преобразуется) или, как теперь принято говорить, кодируется, рецептором в нервный импульс и в таком виде по чувствительным волокнам направляется в ЦНС. Здесь эта информация перерабатывается, отбирается и передается на двигательные нервные клетки, которые посылают нервные импульсы к рабочим органам - мышцам, железам и вызывают тот или иной приспособительный акт - движение или секрецию.
Во время ответной реакции возбуждаются рецепторы рабочего органа и от них в ЦНС поступают импульсы - информация о достигнутом результате. Живой организм, как любая саморегулирующаяся система, работает по принципу обратной связи. Афферентные импульсы, осуществляющие обратную связь, либо усиливают и уточняют реакцию, если она не достигла цели, либо прекращают ее. Таким образом, рефлекс осуществляется не рефлекторной дугой, а рефлекторным кольцом (П. К. Анохин); рефлекс заканчивается по достижении результата.
Рефлекс обеспечивает тонкое, точное и совершенное уравновешивание организма с окружающей средой, а также контроль и регуляцию функций внутри организма. В этом его биологическое значение. Рефлекс является функциональной единицей нервной деятельности.
Вся нервная деятельность складывается из рефлексов различной степени сложности, т. е. является отраженной, вызванной внешним поводом, внешним толчком. Рефлекторный принцип нервной деятельности был открыт великим французским философом, физиком и математиком Рене Декартом в XVII веке.
Развитие рефлекторная теория получила в фундаментальных трудах русских ученых И. М. Сеченова и И. П. Павлова. В 1863 г. в книге "Рефлексы головного мозга" И. М. Сеченов высказал мысль, что не только спинной мозг, как полагал Декарт, но и головной мозг работает по принципу рефлекса: "...без внешнего чувственного раздражения невозможна хоть на миг психическая деятельность г ее выражение - мышечное движение".
И. М. Сеченов писал: "... если выключить все рецепторы, то человек должен заснуть мертвым сном и никогда не проснуться." Это теоретическое положение нашло свое обоснование в клинической практике. С. П. Боткин наблюдал больного, у которого из всех рецепторов тела функционировали один глаз и одно ухо. Как только больному закрывали глаз и затыкали ухо, он засыпал.
В опытах В. С. Галкина собаки, у которых путем операции одновременно были выключены зрительные, слуховые и обонятельные рецепторы, спали по 20 - 23 ч в сутки. Пробуждались они только под влиянием внутренних потребностей или энергичного воздействия на кожные рецепторы. Следовательно, ЦНС работает по принципу рефлекса отражения, по принципу стимул - реакция.
И. П. Павлов открыл условные рефлексы - качественно новую, высшую форму нервной деятельности, свойственную головному мозгу. Он создал рефлекторную теорию в ее современном виде.
Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса. Если лапку лягушки опустить в слабый раствор серной кислоты, возникнет оборонительный рефлекс - лапка отдернется. Однако если снять кожу и тем самым удалить кожные рецепторы, то серная кислота не окажет действия.
То же самое можно наблюдать при разрушении любого другого звена: ЦНС, чувствительных или двигательных нервных волокон. Самое сильное раздражение не вызовет ответной реакции, нервная деятельность будет отсутствовать.
Этим широко пользуются хирурги, применяя во время операции новокаин для анестезии периферических нервов или ганглиоблокатор, прерывающий проведение возбуждения в синапсах. Наркотические вещества центрального действия выключают функцию нейронов ЦНС.
Время рефлекса. Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса (латентный период). Оно слагается из времени, необходимого для возбуждения рецепторов, проведения возбуждения по чувствительным волокнам, ЦНС, двигательным волокнам, и, наконец, скрытого (латентного) периода возбуждения рабочего органа. Большая часть времени уходит на проведение возбуждения через нервные центры - центральное время рефлекса. Это объясняется тем, что в синапсах ЦНС происходит замедление проведения возбуждения, так называемая синаптическая задержка. Чем меньше нейронов входит в состав рефлекторной дуги, тем короче время рефлекса. Поэтому сухожильные рефлексы, возникающие при растяжении сухожилия, имеющие двухнейронную дугу, наиболее быстрые. Их время составляет всего 19 - 23 мс, тогда как время рефлекса моргания, возникающего при раздражении глаза, равно 50 - 200 мс. Наибольшим является время вегетативных рефлексов.
Время рефлекса зависит от силы раздражения и возбудимости ЦНС. При сильном раздражении оно короче, при снижении возбудимости, вызванном, например, утомлением, время рефлекса увеличивается, при повышении возбудимости значительно уменьшается.
Рецептивное поле рефлекса. Каждый рефлекс можно вызвать только с определенного рецептивного поля. Анатомическая область, при раздражении которой вызывается данный рефлекс, носит название рецептивного поля рефлекса. Например, рефлекс сосания возникает при раздражении губ ребенка, рефлекс сужения зрачка - при освещении сетчатки, коленный рефлекс (разгибание голени) - при легком ударе по сухожилию ниже надколенника (рис. 105).

Рис. 105. Методика воспроизведения проприоцептивных рефлексов и
схема рефлекторной дуги коленного рефлекса
Нервный центр. Каждый рефлекс имеет свою локализацию в ЦНС, т. е. тот ее участок, который необходим для его осуществления. Например, центр мочеиспускания находится в крестцовом отделе спинного мозга, центр коленного рефлекса - в поясничном, центр расширения зрачка - в верхнем грудном сегменте спинного мозга. При разрушении соответствующего участка рефлекс отсутствует. Однако выяснилось, что для регуляции рефлекса, его точности недостаточно первичного, или главного, центра, а необходимо участие и высших отделов ЦНС, включая кору большого мозга.
Только при целостности ЦНС сохраняется совершенство нервной деятельности. Нервным центром называется совокупность нервных клеток, расположенных в различных отделах ЦНС, необходимая для осуществления рефлекса и достаточная для его регуляции. Так, если у животного удалить кору полушарий большого мозга, то дыхание сохраняется, так как первичный дыхательный центр находится в продолговатом мозге. Однако во время работы не будет точного соответствия вентиляции легких потребностям организма в кислороде, так как для тонкой регуляции деятельности дыхательного центра необходим не только ствол мозга, но и кора больших полушарий.
Классификация рефлексов. Различают следующие виды рефлексов.
1. По биологическому значению рефлексы подразделяются на пищевые, оборонительные, ориентировочныеи(ознакомление с изменяющимися условиями среды), половые (продолжение рода).
2. По роду рецепторов, с которых они возникают, рефлексы делятся на экстероцептивные, возникающие с рецепторов, воспринимающих раздражения из внешней среды: световые, звуковые, вкусовые, тактильные и др.; интероцептивные, возникающие с рецепторов внутренних органов: механо-, термо-, осмо- и хеморецепторов сосудов и внутренних органов, и проприоцептивные - с рецепторов, находящихся в мышцах, сухожилиях, связках.
3. В зависимости от рабочего органа, участвующего в ответной реакции, рефлексы подразделяются на двигательные, секреторные, сосудистые.
4. По местонахождению главного нервного центра, необходимого для осуществления рефлекса, они делятся на спинальные, например мочеиспускание, дефекация; бульбарные (продолговатый мозг): кашель, чиханье, рвота; мезэнцефальные (средний мозг) : выпрямление тела, ходьба; диэнцефальные (промежуточный мозг) - терморегуляторные; корковые - условные рефлексы.
5. В зависимости от продолжительности различают фазные и тонические рефлексы. Тонические рефлексы длительные, продолжаются часами, например рефлекс стояния. Любое животное может стоять часами благодаря длительному сокращению мышц. Все позные рефлексы относятся к тоническим. Они фиксируют определенное положение тела, а на их фоне разыгрываются другие, короткие, фазные рефлексы, обеспечивающие все виды рабочих, спортивных и других движений.
6. По сложности рефлексы можно разделить на простые и сложные. Расширение зрачка в ответ на затемнение глаза, разгибание ноги в ответ на легкий удар по сухожилию - это простые рефлексы. Примерами сложных рефлексов служат регуляция сердечно-сосудистой системы, процесс пищеварения. В этих случаях конец одного рефлекса служит раздражителем для возникновения другого. Возникают так называемые цепные рефлексы, протекание которых очень демонстративно можно проследить на примере процесса пищеварения. Произвольное проталкивание комка пищи к задней стенке глотки вызывает раздражение ее рецепторов - возникает рефлекс глотания. Пища попадает в пищевод и вызывает его сокращение, продвигающее пищевой комок ко входу в желудок. Раздражение нижней части пищевода приводит к открытию кардинального жома желудка и поступлению пищи в желудок, а последнее вызывает отделение желудочного сока и т. д. Весь процесс пищеварения - сложная цепь рефлексов.
7. По принципу эффекторной иннервации рефлексы можно разделить на скелетно-моторные, или соматические (обеспечивающие двигательные акты скелетной мускулатуры), и вегетативные (функции внутренних органов).
8. В зависимости от того, являются ли рефлексы врожденными или приобретенными в процессе индивидуальной жизни, И. П. Павлов подразделял их на безусловные (врожденные) и условные (приобретенные).
Механизм передачи возбуждения в синапсах. Нервные клетки, образующие рефлекторные дуги, соединяются между собой посредством контактов - синапсов, в которых происходит передача возбуждения от одного нейрона к другому. Синапсы находятся на теле нервной клетки, на дендритах, у периферических окончаний аксона. На каждом нейроне тысячи синапсов, причем большинство - на дендритах (рис. 106).

Рис. 106. Синаптические бляшки (1) окончаний пресинаптических
аксонов образуют соединения на дендритах (2) и теле (3) нейрона
[Стерки П., 1984]
Синапсы по механизму передачи возбуждения разделяются на химические и электрические. Последние находятся в сердечной мышце, гладких мышцах и железистой ткани; в ЦНС наличие их только предполагается.
Синапс, с химической передачей, состоит из синаптической бляшки, пресинаптической мембраны, синаптической щели шириной 30 нм и постсинаптической мембраны (рис. 107).

Рис. 107. Межнейронный синапс [Стерки П., 1984]. 1 - синаптические
пузырьки; 2 - синаптическая цель; 3 - постсинаптические рецепторы; 4
- постсинаптическая мембрана; 5 - синаптическая бляшка; 6 -
митохондрия
В синаптической бляшке медиатор хранится в мелких пузырьках, которых около 3 млн. Под действием нервного импульса наступает деполяризация окончаний аксона, что вызывает повышение концентрации Ca 2+ в нем, и содержимое синаптических пузырьков выбрасывается в синаптическую щель. Роль пускового механизма в выделении медиатора играет повышение концентрации Ca 2+ . Медиатор диффундирует через синаптическую щель и связывается с рецепторными белками постсинаптической мембраны, вызывая в ней возникновение либо возбуждающего постсинаптического потенциала (ВПСП), либо тормозного постсинаптического потенциала (ТПСП).
Медиаторами, вызывающими в нейронах возбуждение, являются ацетилхолин, норадреналин, серотонин, дофамин. Торможение в нейроне вызывает тормозной медиатор - гамма-аминомасляная кислота.
В электрических синапсах синаптическая щель очень узкая (1 - 2 нм), ее пересекают каналы, сквозь которые ионы легко передаются к постсинаптической мембране. Потенциал действия беспрепятственно, без задержки, проводится с одной клетки на другую. Здесь нет химического медиатора; проведение возбуждения по механизму сходно с проведением по нервному волокну.
Особенности нервных центров. Характерными особенностями нервных центров, отличающими их от нервных волокон, являются быстрая утомляемость, очень высокий обмен веществ, т. е. высокая потребность в кислороде и питательных веществах, и избирательная чувствительность к некоторым ядам. Вследствие этих особенностей нарушения кровоснабжения и изменения температуры тела прежде всего сказываются на функции ЦНС: остановка кровоснабжения мозга на 20 с вызывает обморок - потерю сознания; повышение температуры тела до 40 - 42°С - бред, нарушение сознания. Реанимация возможна, если клиническая смерть (остановка сердца и дыхания) продолжалась не более 5 - 6 мин. По истечении большего срока можно восстановить деятельность сердца и даже дыхание, но орган сознания - кора больших полушарий, наиболее чувствительная к изменениям внутренней среды организма, функционировать не будет.
Физиология центральной нервной системы (ЦНС).
ЦНС – система, осуществляющая регуляцию практически всех функций в организме. ЦНС осуществляет связь в единое целое всех клеток и органов нашего организма. С ее помощью происходят наиболее адекватные изменения работы различных органов, направленные на обеспечение той или иной его деятельности. Кроме того, ЦНС осуществляет связь организма с внешней средой, путем анализа и синтеза поступающей к ней информации от рецепторов и формирует ответную реакцию, направленную на поддержание гомеостаза.
Строение ЦНС.
Структурной и функциональной единицей нервной системы является нервная клетка (нейрон). Нейрон - специализированная клетка, способная принимать, кодировать, передавать и хранить информацию, организовывать ответные реакции организма на раздражения, устанавливать контакты с другими нейронами.
Нейрон состоит из тела (сомы) и отростков - многочисленных дендритов и одного аксона (рис1).
Рис.1. Строение нейрона.
Дендриты обычно сильно ветвятся и образуют множество синапсов с другими нервными клетками, что определяет их ведущую роль в восприятии нейроном информации. Аксон начинается от тела клетки аксонным холмиком, функцией которого является генерация нервного импульса, который по аксону проводится к другим клеткам. Длина аксона может достигать одного метра и более. Аксон сильно ветвится, образуя множество коллатералей (паралелльных путей) и терминалей. Терминаль – окончание аксона, с помощью которого образуется синапс с другой клеткой. В ЦНС терминали формируют нейро-нейрональные синапсы, на периферии (за пределами ЦНС) аксоны образуют либо нейро-мышечные, либо нейросекреторные синапсы. Окончание аксона чаще называют не терминалью, а синаптической бляжкой (или синаптической пуговкой). Синаптическая бляшка – это концевое (терминальное) утолщение аксона, служащее для депонирования медиатора (смотрите лекции по синапсу). Мембрана окончаний содержит большое число потенциалозависимых кальциевых каналов, через которые ионы кальция поступают внутрь окончания при его возбуждении.
В большинстве центральных нейронов (т.е. нейронов ЦНС) ПД первично возникает в области мембраны аксонного холмика, и отсюда возбуждение распространяется по аксону к синаптической бляшке. Таким образом, уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний – синапсов.
Каждый нейрон выполняет 2 основные функции: проводит импульсацию и обрабатывает импульсацию (смотрите далее «трансформация ритма возбуждения»). Любой участок нейрона обладает проводимостью. Проведение импульсации (информации) от одной клетки к другой нейрон осуществляет благодаря своим отросткам: аксону и дендритам. Каждый нейрон имеет один аксон и множество дендритов.
Обработка импульсации (обработка информации, трансформация импульсации) - это наиболее значимая функция нейрона, которая осуществляется на аксонном холмике.
Помимо нейронов в ЦНС имеются глиальные клетки, занимающие половину объема мозга. Периферические аксоны (периферические – значит находящиеся за пределами ЦНС) также окружены оболочкой из глиальных клеток. Они способны к делению в течение всей жизни. Размеры 3-4 раза меньше, чем нейроны. С возрастом их число увеличивается.
Функции клеток глии многообразны:
1) они являются для нейронов опорным, защитным и трофическим аппаратом;
2) поддерживают определенную концентрацию ионов кальция и калия в межклеточном пространстве;
3) активно поглощают нейромедиаторы, ограничивая, таким образом, время их действия.
Классификация нейронов
Зависимости от отделов ЦНС: вегетативные и соматические
По виду медиатора, которая выделяется окончаниями нейрона: адренэргические(НА) и т.д
По влиянию бывают возбуждающие и тормозящие
По специфичности воспринимающей сенсорной информации нейроны высших отделов ЦНС бывают моно и полимодальные
По активности нейронов бывают:фоноактивные, молчащие- которые возбуждаются только в ответ на раздражение.
По источнику или направении передачи информации: афферентные, вставочные, эфферентные
Рефлекторный принцип деятельности ЦНС.
Основным механизмом деятельности ЦНС является рефлекс. Рефлекс - это ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС. Например, отдергивание руки при уколе, смыкание век при раздражении роговицы – это тоже рефлекс. Отделение желудочного сока при попадании пищи в желудок, дефекация при наполнении прямой кишки, покраснения кожи при тепловом воздействии, коленный, локтевой, Бабинского, Розенталя – это все примеры рефлексов. Количество рефлексов безгранично. Общим для них всех является обязательное участие в их реализации ЦНС.
Другим определением рефлекса, также подчеркивающим роль ЦНС, является следующее: рефлекс –это центробежный ответ на центростремительное раздражение. (В приведенных примерах самостоятельно определите, что является центробежным ответом, а что раздражением. Раздражение всегда центростремительное, т.е. действующий на рецепторы раздражитель вызывает импульсацию, которая поступает в ЦНС).
Структурной основой рефлекса, его материальным субстратом является рефлекторная дуга (рис.2).

Рис. 2.Рефлекторная дуга
Рефлекторная дуга состоит из 5 звеньев :
1) рецептора;
2) афферентного (чувствительного, центростремительного) звена;
3) вставочного звена (центрального);
4) эфферентного (двигательного, центробежного) звена;
5) эффектора (рабочего органа).
Участок тела, содержащий рецепторы, при раздражении которых возникает определенный рефлекс, называется рецептивным полем рефлекса.
Рефлекс может осуществляться только тогда, когда сохранена целостность всех звеньев рефлекторной дуги.
Нервный центр.
Нервный центр (центр ЦНС или ядро) – это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса. Т.е. каждый рефлекс имеет свой центр: существует центр коленного рефлекса, свой центр у локтевого рефлекса, свой - у мигательного, есть сердечно-сосудистый, дыхательный, пищевой центры, центры сна и бодрствования, голода и жажды и т.д. В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС, т.е. сложное объединение большого количества центров.
Объединение нервных центров (ядер) между собой осуществляется проводящими путями ЦНС с помощью нейро-нейрональных (межнейронных) синапсов. Существует 3 типа соединения нейронов: последовательное, дивергентное и конвергентное.
Нервные центры обладают рядом характерных функциональных свойств, которые во многом обусловлены этими тремя типами нейронных сетей, а также свойствами межнейронных синапсов.
Основные свойства нервных центров:
1. Конвергенция (схождение) (рис.3). В ЦНС к одному нейрону могут сходиться возбуждения от различных источников. Эта способность возбуждений сходиться к одним и тем же промежуточным и конечным нейронам получила название конвергенции возбуждений

Рис.3. Конвергенция возбуждения.
2. Дивергенция (расхождение ) - расхождение импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.

Рис.4. Дивергенция возбуждения.
3. Возбуждение в нервных центрах распространяется односторонне - от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
4. Возбуждение в нервных центрах проводится медленнее , чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапсы (синаптическая задержка), которых в ядре много.
5. В нервных центрах осуществляется суммация возбуждений . Суммация – сложение допороговых импульсов. Различают два вида суммации.
Временная или последовательная , если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях локальные токи на постсинаптической мембране воспринимающего нейрона суммируются и доводят ее деполяризацию до уровня Е к, достаточного для генерации нейроном потенциала действия. Временной данная суммация называется, потому что на нейрон в течение некоторого промежутка времени приходит серия импульсов (раздражений). Последовательной она называется, потому что реализуется в последовательном соединении нейронов.
Пространственная или одновременная - наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы. Пространственной данная суммация называется, потому что раздражитель действует на некоторое пространство рецептивного поля, т.е. несколько (минимум 2) рецепторов разных участков рецептивного поля. (Тогда как временная суммация может реализоваться при действии серии раздражителей на один и тот же рецептор). Одновременной она называется, потому что информация к нейрону приходят одновременно по нескольким (минимум 2) каналам связи, т.е. одновременная суммация, реализуется конвергентным соединением нейронов.
6.Трансформация ритма возбуждения - изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
1) понижающая трансформация , в основе которой, лежит явление суммации возбуждений, когда в ответ на несколько пришедших допороговых возбуждений к нервной клетке, в нейроне возникает только одно пороговое возбуждение;
2) повышающая трансформация , в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения на выходе.
7. Рефлекторное последействие - заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено двумя причинами:
1) длительной следовой деполяризацией мембраны нейрона, на фоне прихода мощной афферентации (сильной чувствительной импульсации), вызывающей выделение большого количества (квантов) медиатора, что обеспечивает возникновение нескольких потенциалов действия на постсинаптической мембране и, соответственно, кратковременное рефлекторное последействие;
2) пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки". Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление. Примером последействия может служить хорошо всем известная жизненная ситуация, когда даже после прекращения действия сильного эмоционального раздражителя (после прекращения ссоры) еще какое-то более или менее продолжительное время продолжается общее возбуждение, артериальное давление остается повышенным, сохраняется гиперемия лица, тремор кистей.
8. Нервные центры обладают высокой чувствительностью к недостатку кислорода. Нервные клетки отличаются интенсивным потреблением О 2 . Мозг человека поглощает около 40-70 мл О 2 в минуту, что составляет 1/4-1/8 часть всего количества О 2 , потребляемого организмом. Потребляя большое количество О 2 , нервные клетки высокочувствительны к его недостатку. Частичное прекращение кровообращения центра ведет к тяжелым расстройствам деятельности его нейронов, а полное прекращение - к гибели в течение 5-6 мин.
9. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических вещест в, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам. Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
10. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми. Это обусловлено резким уменьшением запасов медиатора, уменьшением чувствительности к медиатору постсинаптической мембраны, уменьшением ее энергетических запасов, что наблюдается при длительной работе и является основной причиной развития утомления.
11. Нервные центры, как и синапсы, обладают низкой лабильностью, основной причиной которой является синаптическая задержка. Суммарная синаптическая задержка, наблюдающаяся во всех нейро-нейрональных синапсах при проведении импульсации по ЦНС, или в нервном центре называется центральной задержкой.
12. Нервные центры обладают тонусом , который выражается в том, что даже при отсутствии специальных раздражений, они постоянно посылают импульсы к рабочим органам.
13. Нервные центры обладают пластичностью - способностью изменять собственное функциональное назначение и расширять свои функциональные возможности. Так же пластичность можно определить, как способность одних нейронов брать на себя функцию пораженных нейронов того же центра. Именно, с явлением пластичности связана способность восстанавливать двигательную активность конечностей, например, ног, утраченную в результате травм спинного мозга. Однако это возможно только при поражении части нейронов данного центра или при сохранении целостными части проводящих путей ЦНС. При полном разрыве спинного мозга восстановление двигательной активности оказывается невозможным. Кроме того, нейроны одного центра, например, сгибателей не могут брать на себя функцию нейронов другого центра - разгибателей. Т.е. явление пластичности центров ЦНС ограничено.
14. Окклюзия (запирание ) (рис.5) - это сложение пороговой импульсации. Окклюзия осуществляется (так же как и пространственная суммация) в конвергирующей системе соединения нейронов. Одновременной активации нескольких (минимум двух) рецепторов сильным или сверхсильным раздражителями к одному нейрону будут конвергировать несколько пороговых или сверхпороговых импульса. На этом нейроне будет происходить окклюзия, т.е. эти два раздражителя он ответит с той же максимальной силой, что и на каждый из них отдельности. Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при отдельном раздражении каждого афферентного входа в отдельности.

Рис.6. Явление окклюзии в ЦНС.
Явление окклюзии приводит к снижению силы ответной реакции. Окклюзия имеет охранительное значение, предотвращая перенапряжение нейронов при действии сверхсильных раздражителей.
Похожая информация.